
Placental Mammals are more abundant than Marsupials in almost all modern ecosystems; however in the Late Cretaceous the Metatherans (the wider group that includes the Marsupials - without knowledge of the reproductive system of an ancient Mammal it is impossible to say whether we would consider if a Marsupial or not, so this term is restricted to the last common ancestor of all living Marsupials and everything descended to it, while the term Metatheran is used to indicate anything more closely related to the modern Marsupials than the modern Placentals) were much more widespread and diverse, inhabiting a broad range of ecological niches, even in Northern Hemisphere regions from which they are now absent. Sixty species of Metatherans have been described from the Late Cretaceous of North America to date, although the majority of these are known only from teeth and other fragmentary remains, and intact crania (skulls) are extremely rare. Didelphodon vorax was first described in 1889 by palaeontologist Othniel Charles Marsh. It was a large Mammal by Cretaceous standards, probably reaching around 5 kg in mass, with teeth suggesting a carnivorous diet and a skeleton interpreted as implying a possible semi-aquatic lifestyle.
In a paper published in the journal Nature Communications on 8 December 2016, Gregory Wilson of the Department of Biology and Burke Museum at the University of Washington, Eric Ekdale of the Department of Biology at San Diego State University and the Department of Paleontology at the San Diego Natural History Museum, John Hoganson of the North Dakota Geological Survey, Jonathan Calede, also of the Department of Biology and Burke Museum at the University of Washington and Abby Vander Linden of the Graduate Program in Organismic and Evolutionary Biology at the University of Massachusetts describe four new specimens of Didelphodon vorax, including a nearly complete skull, two maxilae (upper jaws) and a detached snout from the Late Cretaceous Hell Creek Formation of Montana and North Dakota.

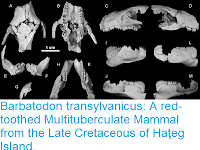
Skull of the Late Cretaceous marsupialiform Didelphodon vorax. Specimen NDGS 431, in anterior (a), left lateral (b), posterior (c), dorsal (d) and ventral (e) views, and specimen UWBM 94084, in anterior (f), left lateral (g), dorsal (h) and ventral (i) views. All images from digitally rendered, micro-CT scans. as, alisphenoid; bo, basioccipital; bs, basisphenoid; C, upper canine; I1–4, upper incisors 1–4; inp, internarial process; inf, incisive foramen; iof, infraorbital foramen; ip, interparietal; ju, jugal; M1–4, upper molars 1–4; mapf, major palatine fenestra; mp, mastoid process; mpf, minor palatine foramen; mx, maxilla; na, nasal; P1–3, upper premolars 1–3; pa, parietal; pe, petrosal; pgp, postglenoid process; pmx, premaxilla; ppt, postpalatine torus; ps, presphenoid; pt, pterygoid; rtpp, rostral tympanic process of the petrosal; so, supraoccipital; sq, squamosal. Scale bar, 10 mm. Wilson et al. (2016).
These cranial remains enable some reconstruction of the biology and ecology of Didelphodon vorax. From the anatomy of the whole cranium (which is not thought to have come from a fully mature individual) Wilson et al. estimate that the living animal has a bite force measured at the canines of 218 Newtons. This is lower than that of many living and fossil Mammals (for example the European Badger has a bite force of 244 Newtons), but, due to the small mass of Didelphodon vorax, translates into a bite force quotient (a ration of bite force to size) of 201, higher than that of any other Mammal for which this has been calculated, living or fossil; exceeding that of the Tasmanian Devil (166), Spotted Hyena (114) and African Lion (116) as well as the extinct Marsupial Lion (193) and Dire Wolf (157).

Reconstruction of the skull and jaw of Didelphodon vorax. The reconstruction is shown in anterior (a), posterior (b), dorsal (c), ventral (d), and right lateral (e) views. It is based on NDGS 431 (basicranium, palate, skull roof and dentition), UWBM 94084 (rostrum), UWBM 94500 (palate and maxillo-jugal contact), SCNHM VMMa 20 (maxillo-jugal contact) and UWBM 102139 (dentary). The areas not preserved in the actual specimens include the lower incisors, some upper incisors, parts of the orbitotemporal and occiput regions, and some sutures. Uncertainties in sutures ar represented by dashed lines. Scale bar, 10 mm. Wilson et al. (2016).
Such a bite force, combined with the dentition of Didelphodon vorax, which is typical of a carnivorous diet, would appear to suggest the species was a durophagous carnivore similar to a modern Hyena or Big Cat. However examination of the wear on the teeth of the specimens does not match that seen on such animals, rather being closer to that seen on malacavorous animals (animals that feed mainly on Molluscs). Since this diet does not match the dentition of Didelphodon vorax, Wilson et al. suggest the species may have been omnivorous in diet.
See also...





Follow Sciency Thoughts on Facebook.